牛乳中脂肪酶活力檢測方法與影響因素研究
2021-08-06 15:53:13 來源: 世界杯賽程預測 導刊
陳柏錫,黃豔豔,曾燕萍,李 斌,鄒蘭鈺,彭遠霞,張 偉
(四川新希望乳業有限公司洪雅陽平分公司,四川眉山 620000)
摘 要:原料乳中的某些嗜冷菌會在代謝過程中產生耐熱脂肪酶,較高的脂肪酶活力會破壞牛乳中的脂肪球結構並產生多種脂肪酸,遊離的脂肪酸易被氧化,產生異味。本文闡述一種利用脂肪酶分解底物產生具有特定吸收波長的物質實現準確檢測牛乳中脂肪酶活力的方法,探究了原料奶的運輸時間、脫冷儲運對脂肪酶活力的影響,以及脂肪酶活力對UHT滅菌乳風味與貨架期的影響,期望為相關人員提供一定的參考依據。(四川新希望乳業有限公司洪雅陽平分公司,四川眉山 620000)
關鍵詞:乳脂肪酶活力;比色法檢測;原奶儲運;UHT滅菌乳
超高溫滅菌乳(Ultra High Temperature,UHT乳)經過短時高溫的滅菌後,產品達到商業無菌的狀態,因此UHT乳具有保質期長、可常溫儲運等特點[1]。但超高溫滅菌乳在儲存過程中會出現風味異常、脂肪上浮等問題,嚴重影響產品的品質和貨架期,該現象的主要原因是由UHT乳中脂肪酶導致的。牛乳中的脂肪酶主要有兩種來源,一種是天然存在於生乳中,稱為天然脂肪酶;另一種是牛乳中的微生物代謝所產生,稱為微生物脂肪酶[2]。牛乳中的微生物脂肪酶主要來源於嗜冷菌,雖然嗜冷菌本身的繁殖不會嚴重影響牛乳的穩定性,且在較為溫和的熱處理中(如巴氏殺菌)也容易將其殺死,但某些嗜冷菌的代謝產物是耐熱脂肪酶,耐熱脂肪酶在經過巴氏殺菌甚至超高溫滅菌後,仍不能被完全滅活。殘留的脂肪酶會分解牛乳中的脂肪,造成脂肪上浮和風味異常,嚴重破壞滅菌乳的口感和風味,導致滅菌乳的貨架期大幅縮短[3]。所以需要一種能夠快速、準確且價廉的檢測方法檢測原料乳中脂肪酶的活力以及探究牛乳中脂肪酶活力的變化規律。
4-硝基苯基辛酸酯(PNP-C)在脂肪酶的作用下會產生在316 nm處有吸光度的物質,根據比爾-朗博定律,在一定濃度下,吸光度大小與吸光度物質濃度呈正比,實驗在底物濃度相同的情況下,產生吸光物質濃度與脂肪酶活力成正比。本實驗使用脂肪酶標準品配製不同濃度的脂肪酶溶液,分解底物產生吸光度後標定曲線,以此測定樣品中脂肪酶活力。
1 材料與方法
1.1 材料與儀器
1.1.1 材料
樣品來源於HN、ZX、ZN和XN共4家牧場生乳;
5 U/mL脂肪酶溶液、5 mmol/L4-硝基苯基辛酸酯底物溶液、50 mmol/L pH7.2磷酸緩衝液、終止液:將乙腈與甲酸按照12∶1的比例混勻即可作為終止液。
1.1.2 儀器
紫外分光光度計、水浴加熱器、冷凍離心機、移液槍、移液管、水相濾頭(0.22 μm)等。
1.2 實驗步驟
將5 U/mL脂肪酶溶液配製成100 mU/mL的脂肪酶稀釋液備用,分裝好的底物在室溫下用流水解凍。取20 mL原料奶於50 mL離心管中,於10 000 r/min離心脫脂15 min。離心後將上層脂肪撇去,吸取100 μL清液於900 μL磷酸緩衝液中,混勻,製成1 mL樣品處理液。分別吸取300 μL樣品處理液於兩個2 mL離心管中,其中一個為加標樣,並向兩個離心管中分別加入180 μL和120 μL磷酸緩衝液,並向加標樣的離心管中加入60 μL 100 mU/mL脂肪酶稀釋液,最後向離心管中各加入120 μL底物溶液,製成待測樣品和加標樣品。
製備標準曲線:取4個2 mL離心管,每個離心管加入480 μL、450 μL、420 μL和360 μL磷酸緩衝液,向離心管中加入120 μL底物和0 μL、30 μL、60 μL和120 μL脂肪酶稀釋液,混勻後即製成0 mU/mL、5 mU/mL、10 mU/mL和20 mU/mL的標準曲線。將樣品和標準曲線置於44 ℃水浴加熱1 h取出,向每個離心管中加入1.3 mL終止液,搖勻,即可上機檢測。
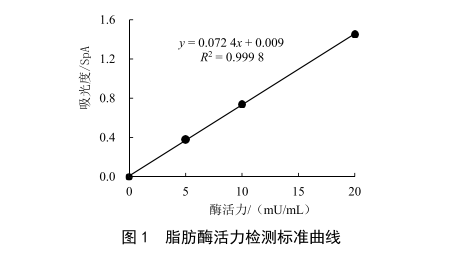
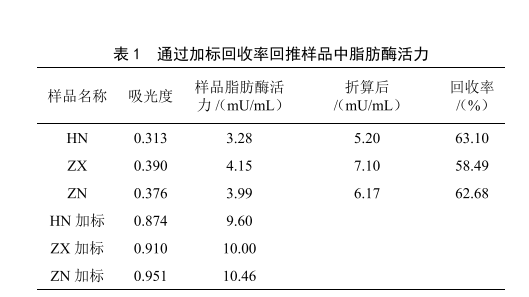
1.3 標準曲線與數據處理
標準曲線見圖1,樣品中脂肪酶活力見表1。由HN、ZX、ZN 3批原料奶的吸光度計算出樣品中的脂肪酶活力,再根據加標回收率折算出樣品中脂肪酶活力準確值。需注意的是,樣品的回收率應在50%以上(如表1),當回收率偏低時,有可能是離心效果不佳,導致脂肪剩餘過多導致的,其原因是樣品中的脂肪酶不僅會與底物反應,還會與牛乳中剩餘的脂肪反應,在相同的反應時間內,剩餘脂肪的含量越高,導致產生吸光物質的濃度比理論值越低,影響檢測結果。原料乳離心脫脂率通常為90%~95%。為補償剩餘脂肪產生的影響,每個樣品必須進行平行試驗和加標回收。
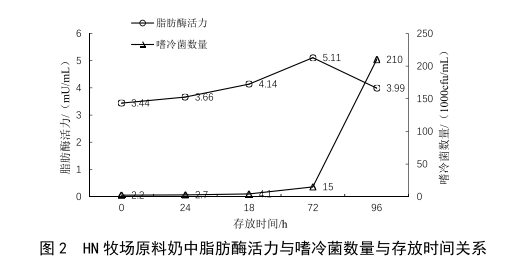
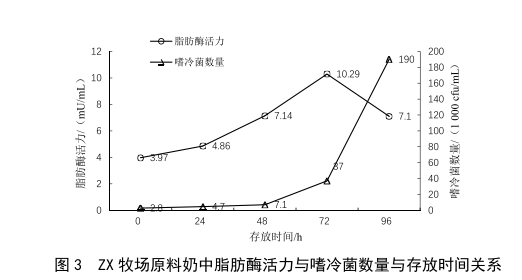
2.1 原奶儲運時間對脂肪酶活力的影響
牛乳中耐熱脂肪酶是部分嗜冷菌的代謝產物,在冷藏條件下,隨著嗜冷菌的繁殖與代謝,牛乳中的脂肪酶活力應隨之升高。本次實驗取兩個不同牧場的原料奶各5份,置於2~6 ℃條件下冷藏避光保存,每隔24 h對其脂肪酶活力和嗜冷菌數量進行檢測,繪製出的結果見圖2、
圖3。
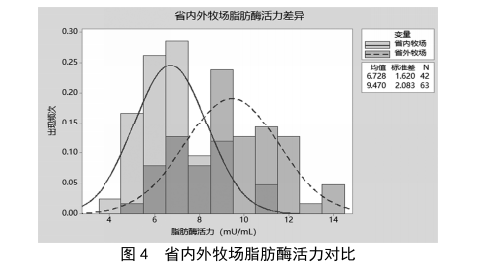
在乳品生產加工的供應鏈體係中,原料奶的運輸時長會隨著牧場與工廠距離的增加而增加,跨省牧場的運輸時間在20 h以上,運輸溫度均在4 ℃,原奶運輸過程可近似理解為低溫儲存的過程,本次實驗對省外牧場與省內牧場的原料奶進行脂肪酶檢測,並統計其數據,結果見圖4。由圖4可知,省外牧場與省內牧場的原料奶中脂肪酶活力均基本呈現正態分布,省外牧場的脂肪酶活力多數在8~11 mU/mL(平均值9.47 mU/mL),省內牧場的脂肪酶活力普遍在5~8 mU/mL(平均值6.73 mU/mL),省外牧場奶源的脂肪酶活力明顯高於省內牧場奶源,與原奶冷藏實驗得到的結論一致。
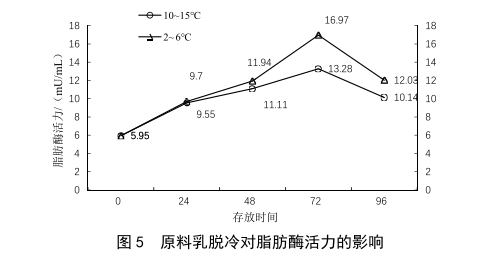
奶車運輸和原奶在工廠的奶倉中儲存均在4 ℃左右,當奶車運輸或原奶儲存出現脫冷時(10~15 ℃),也會對原料奶的質量造成影響[4],取相同批次的XN牧場原料乳,分別在2~6 ℃與10~15 ℃環境中保存,模擬奶車正常運輸與脫冷的狀態,其脂肪酶檢測結果如圖5所示。當運輸或儲存出現脫冷時,脂肪酶活力上升的速度也明顯增加。在最高點時(72 h),脫冷條件下的脂肪酶活力也明顯高於冷藏條件。
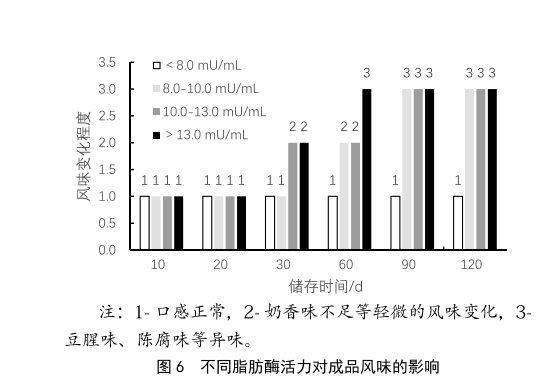
UHT產品由於其經過了超高溫滅菌工藝,經過無菌灌裝後,產品內部實現商業無菌狀態,因此UHT產品可在常溫下儲存,且保質期較長,通常可達到6個月。但UHT產品在儲存時經常會出現脂肪上浮、風味異常、口感變差等情況,這類情況會嚴重影響產品的貨架期,並影響消費者的購買體驗。原料奶中產生的耐熱脂肪酶在經過超高溫滅菌後,仍然不能被完全滅活,這部分脂肪酶會持續分解乳脂肪,產生脂肪酸,引起脂肪上浮或氧化味[5]。本次實驗選取不同梯度的脂肪酶活力的原料奶,將其對應的UHT成品進行常溫儲存,模擬貨架陳列。感官品評小組在10 d、20 d、30 d、60 d、90d和120 d後進行口感品評,客觀的記錄其風味,其感官結果見圖6。
3 結論
牛乳中嗜冷菌代謝的耐熱脂肪酶不會被超高溫滅菌工藝完全滅活,殘留的脂肪酶會持續分解成品乳中的脂肪,縮短產品貨架期[6]。經實驗表明,原料奶中脂肪酶活力越高,對應成品的風味變化越快,越容易產生脂肪氧化的豆腥味或陳腐味。原料奶中的脂肪酶活力隨著儲運時長的增加而不斷提高,在72 h後降低,即使在輕微脫冷的情況下,脂肪酶活力的上升速度也會明顯提高,所以在供應鏈係統中縮短運輸、儲存時間是控製脂肪酶活力的有效手段,對奶車的運輸和原奶的儲存溫度的監控也至關重要。另外,供應鏈上遊的奶源基地的衛生條件直接影響了牛乳中的微生物指標,奶源基地良好的衛生條件是保證原料奶中較低嗜冷菌數量和較低脂肪酶活力的必要條件。
參考文獻
[1]張甦.哈爾濱地區生鮮牛乳中嗜冷菌數量與蛋白酶酶活力關係的測定[J].中國乳品工業,2012,40(7):41-42.
[2]史玉東,康小紅.淺析UHT乳中耐熱酶與產品質量問題的關係[J].農產品加工(學刊),2008(2):79-81.
[3]王歡,何臘平,周換景,等.脂肪酶活力測定方法及其在篩選產脂肪酶微生物中的應用[J].生物技術通報,2013(1):203-208.
[4]張愛霞.耐熱酶與乳成分對UHT乳穩定性影響的研究[D].保定:河北農業大學,2003.
[5]郭德軍,孫愛民,紀振傑.原料乳中嗜冷菌及其主要熱穩定性酶類的研究進展[J].中國乳品工業,2006(8):46-49.
[6]胥伯強.嗜冷菌對長貨架期乳製品的影響[J].中國食品工業,2003(3):50.






熱點推薦
- 世界杯2022赛程时间表最新
-

全十紅紅稗餅幹|中秋團圓,回家必備
-

和汪氏蜂蜜共同來普及蜂蜜結晶現象
-

拿坡海開啟西餐加盟新潮流, 大眾化家庭小西餐成未來新趨勢
-

摩購空間“霸王餐”來襲 打破傳統營銷套路帶動商家業績翻倍
-
牛乳中脂肪酶活力檢測方法與影響因素研究
-

果蔬中農藥殘留快速檢測的質量控製探討
-

氣相色譜-質譜聯用法測定綠茶中4種菊酯類殺蟲劑農藥殘留量
-

幾種常見食品中亞硝酸鹽含量的測定
-
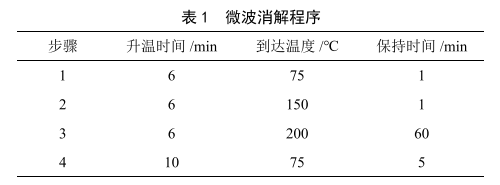
微波消解-電感耦合等離子體質譜法測定大米中的砷、鉛、鎘、鉻
-
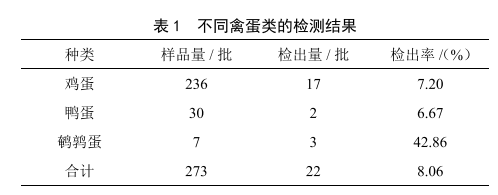
2019—2020年北海市禽蛋中6種獸藥殘留檢測結果分析
-
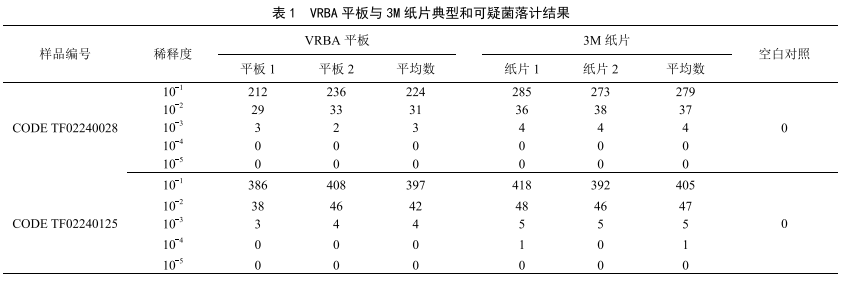
食品中大腸菌群檢測結果與分析
-
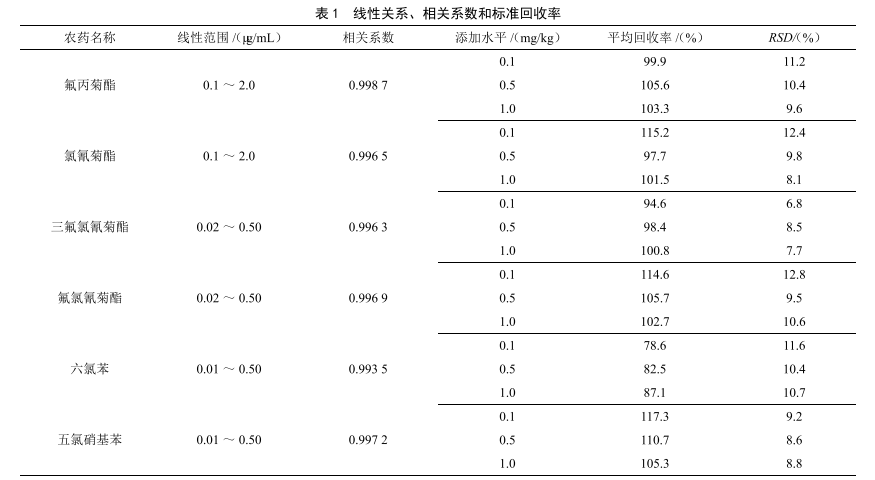
氣相色譜法測定大米中多種農藥殘留量
-
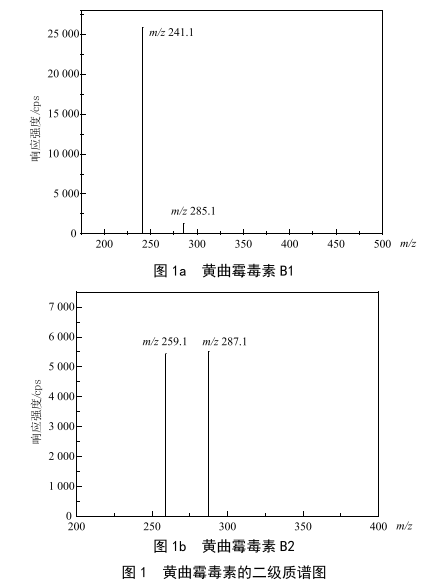
分散液液微萃取-超高效液相色譜-串聯質譜法測定茶飲料中黃曲黴毒
-

辣椒及其製品中辣椒素含量檢測及辣度分級
-
火焰原子吸收光譜法測定食品中鋅的方法驗證
-
對硫磷等6種農藥殘留熒光免疫檢測芯片的研發及應用
-

山東省濰坊市市售蔬菜中農藥殘留現況及慢性膳食暴露風險評估
-

食品檢驗檢測機構質量控製的探討
-
快速檢測技術在世界杯賽程預測 中的應用研究進展
-
基於X射線圖像的堅果盒異物檢測
-
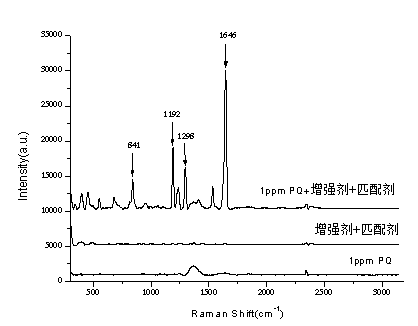
表麵增強拉曼光譜法快速檢測調味品中的百草枯
-
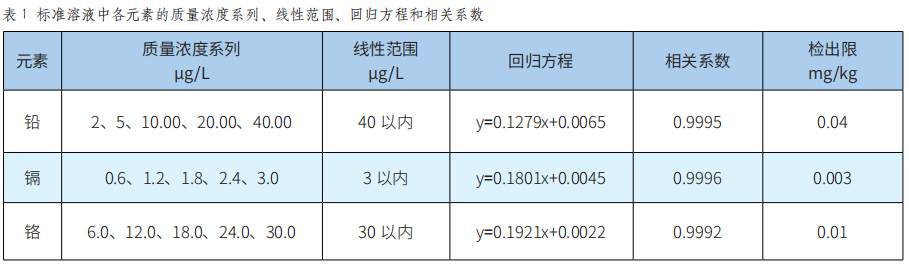
潞黨參重金屬汙染物含量的評估
-
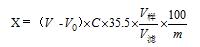
電位滴定法測定特殊醫學用途食品中氯化物的不確定度評定
-
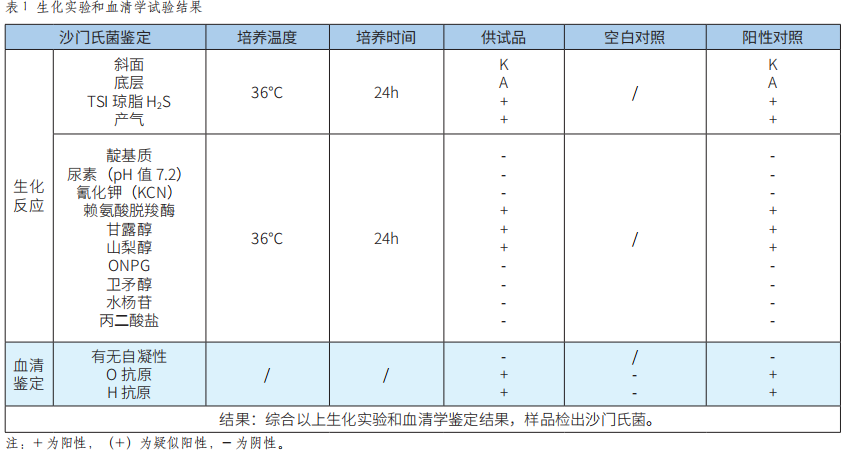
API 20E鑒定係統等兩種檢測方法應用於食品中沙門氏菌的檢測結果
-

2020年上海市青浦區肉製品中單增李斯特菌分子分型及耐藥性分析
-
一種基於ATP熒光反應的潔淨度檢測係統的開發與驗證
-
固相萃取-高效液相色譜-二極管陣列檢測法測定飲料中9種人工合成著
-

一種檢測鹽黴素膠體金試紙條的研製及其應用
-

高效液相色譜法測定綠豆糕中5種常見的食品添加劑