氨基甲酸乙酯的研究進展
2020-07-28 13:34:58 來源: 世界杯賽程預測 導刊
氨基甲酸乙酯的研究進展
氨基甲酸乙酯(EC)是一種廣泛存在於發酵食品和酒精飲料中的化學物質,被國際癌症研究機構(IARC)列為2A類致癌物。新的證據表明,長期接觸EC可能會導致神經係統紊亂。因此,人們對食品中EC的形成機理及其毒性機製進行了廣泛的研究。由於EC對人類健康存在潛在威脅,人們通過物理、化學、酶等方法來緩解食品中的EC。天然產物可以通過調節氧化應激來防止EC引起的毒性。本文綜述了EC的形成機理與 EC對人體各器官的毒性作用以及目前消減EC的策略進展。
關鍵詞:氨基甲酸乙酯;毒性機製;消減策略
引言
氨基甲酸乙酯(Ethyl carbamate,EC)是一種在發酵過程中自發形成的化學物質。乳酪、麵包、酸奶、葡萄酒、威士忌、醬油等發酵食品和酒精飲料中都含有EC。一項體外研究表明,EC具有抑製細菌、植物組織和大鼠癌生長的潛力[1]。
在進口發酵酒精飲料中檢測到相對較高水平的EC時,發酵飲料中EC首先引起了人們的注意[2]。盡管分析方法和條件的多樣性阻礙了不同發酵飲料中EC的比較,但一些調查得出了類似的結論,例如,加拿大和韓國的發酵飲料都顯示白蘭地中EC含量最高[3]。中國黃酒中的EC含量是其他酒精飲料的近兩倍[4]。
EC的形成機理
形成EC的前體物質包括尿素、瓜氨酸、磷酸氨基甲酰、氰化物和碳酸二乙酯。發酵過程中,溫度、酸度、微生物性質等因素都會影響EC的產生[5]。根據所考慮的食品或飲料的性質、它們的生產過程以及原材料中可獲得的EC前體,有許多種自然形成EC的方法。在酸性介質中尿素與乙醇反應是製備EC最常用的方法之一。
第二個途徑是尿素熱分解成氨和氰酸,並通過氰酸和乙醇的反應生成EC[6]。尿素在牛奶等原料中含量不可忽略或在食品(或飲料)加工過程中形成[7]。
最後是通過氰化物陰離子,植物可以產生氰基糖苷和通常相應的水解酶。因此氰基糖苷能產生糖和氰醇後迅速分解成氰化氫。例如,作為熱帶地區第三大食物來源的木薯含有大量的氰化物,可能導致氰化物中毒[8]。
EC的毒性機製
自從Anderson等人[9]首次證明EC可誘發小鼠肺癌以來,EC在不同組織中的致癌性機製一直是研究的主題(表1)。最主要的途徑是在肝微粒體酯酶的幫助下分解90%以上的EC,並產生乙醇、氨和二氧化碳。在細胞色素P450(CY2E1)催化下,不到0.5%的EC被水解成氨基甲酸乙烯酯(VC)。上述相同的酶將VC轉化為VC環氧化合物(VCO),最後強親電的VCO與腺苷反應,生成1-N6乙烯基腺苷加合物。也有報道描述了EC誘導姐妹染色單體交換[10]。

EC的消減策略
原料改性與發酵工藝優化
尿素是EC的主要前驅體,通過還原尿素可以減少EC的生成。大米加工前的洗滌、拋光、預處理等精製工藝可以降低尿素的含量[14]。從氰化物中生成EC是飲料的主要途徑。通過減少光照,縮短貯藏時間,可以防止氰化物的釋放。加工和儲存過程中的高溫條件會增加EC含量。因此,降低溫度是抑製EC形成的有效物理技術。優化條件可以減少EC的生成。
發酵微生物的改性
通過兩種途徑來實現:第一,精氨酸酶基因的缺失和沉默在許多研究中都得到了重視,並且這些嚐試都顯示出了良好的效果;第二,與尿素代謝相關的基因表達的增強或抑製也得到了廣泛的研究,許多改性發酵菌在白酒工業發酵中顯示出潛在的應用前景。精氨酸酶是尿素生成的關鍵酶,催化精氨酸的降解,生成尿素和鳥氨酸。用葡萄酒陳釀和儲存的微生物和物理化學條件對EC生產有影響[15]。
添加酸性脲酶
在大鼠胃腸道乳酸菌中首次發現酸性尿素酶,其最適pH值範圍與葡萄酒發酵條件相適應。這種方法的效率似乎隨許多因素而變化,包括葡萄酒的類型、抑製因子的含量和使用條件[16]。通常,已證實的抑製劑是由氟化物、蘋果酸、乙醇和葡萄酒中的酚類化合物組成的。
目前,研究者致力於通過不同的策略來提高尿素的去除效率。分離潛在的替代性尿素酶以去除酒精飲料中的尿素已經成為一個活躍的研究領域。目前商業級酸性脲酶主要來自發酵乳杆菌。雖然酸性脲酶用於去除尿素的可行性已被廣泛證明,酸性脲酶在工業發酵中的大規模應用仍然受到其耗時和成本的限製。
直接添加氨基甲酸乙酯降解酶
氨基甲酸酶是一種能直接降解EC的酶。1990年,從小鼠糞便中分離出氨基甲酸酶,一種檸檬酸杆菌能將EC化學計量分解為氨和乙醇。它被認為對幾種氨基甲酸酯和酰胺類化合物的水解是有效的,但缺乏對尿素、N-烷基脲和有機酸乙酯的水解能力。這種酶在較高濃度的乙醇和酸性條件下是不活躍的[17]。日本研究人員發現了一種利用表達這種新型氨基甲酸酶的宿主細胞降解EC的新方法。利用氨基甲酸酶編碼基因轉化大腸杆菌表達氨基甲酸酶,提高酶活性[18]。
總結
對於未來的研究,在不影響發酵飲料質量或風味的情況下實現經濟節約的方法需要進一步探索。首先,應探索能直接降解EC的氨基甲酸酶的應用。氨聚糖酶的篩選、性質和應用可參考酸性脲酶。第二,NCR中磷酸化的調節在降低EC方麵也有希望。相關的中間體和相應的功能伴隨著不同化合物之間的變化需要得到充分的證明。這些途徑的實現應該啟動對NCR調控因素和其他調控機製的深入研究。第三,優化可能加速EC減排策略商業化的因素。值得注意的是,在探索降低EC的方法時,發酵飲料的安全性不容忽視。
參考文獻
[1] Anderson N, Paul SH, Henry SM. 1943. Induction of pulmonary tumors in mice with ethyl carbamate (urethane). J Natl Cancer Inst 4:309–19.
[2] Conacher HBS, Page BD. 1986. Ethyl carbamate in alcoholic beverages: a Canadian case history. Proceedings of Euro Food T ox II. Switzerland:European Society of T oxicology. p 237–42.
[3] Alexander J, Auounsson G, Benford D, Cockburn A, Cravedi J, Doglitti E.2007. Ethyl carbamate and hydrocyanic acid in food and beverages.Scientific opinion of the panel on contaminants. EFSA J 551:1–44.
[4] W u P, Pan X, W ang L, Shen X, Y ang D. 2012. A survey of ethyl carbamate in fermented foods and beverages from Zhejiang, China. Food Control 23(1):286–8.
[5] Weber J V, Sharypov V I. Ethyl carbamate in foods and beverages: a review[J]. Environmental Chemistry Letters, 2009, 7(3): 233-247.
[6] Schaber PM, Colson J, Higgins S, Thielen D, Anspach B, Brauer J(2004) Thermal decomposition (pyrolysis) of urea in an openreaction vessel. Thermochim Acta 424:131–142
[7] Eicher R, Bouchard E, Tremblay A (1999) Cow level sampling factors affecting analysis and interpretation of milk urea concentrations in 2 dairy herds. Can Vet J 40(7):487–492
[8] Cardoso AP, Mirione E, Ernesto M, Massaza F, Cliff J, Rezaul Haque Mand Bradbury JH (2005) Processing of cassava roots to remove cyanogens. J Food Compost Anal 18(5):451–460
[9] Anderson N, Paul SH, Henry SM. 1943. Induction of pulmonary tumors in mice with ethyl carbamate (urethane). J Natl Cancer Inst 4:309–19.
[10] Roberts GT, Allen JW. 1980. Tissue-specific induction of sister chromatid exchanges by ethyl carbamate in mice. Environ Mol Mutagen 2(1):17–26.
[11] Guengerich FP, Kim DH. 1991. Enzymic oxidation of ethyl carbamate to vinyl carbamate and its role as an intermediate in the formation of 1,N6-ethenoadenosine. Chem Res T oxicol 4(4):413–21.
[12] Sakano K, Oikawa S, Hiraku Y, Kawanishi S. 2002. Metabolism of carcinogenic urethane to nitric oxide is involved in oxidative DNA damage.Free Radical Biol Med 33(5):703–14.
[13] Roberts GT, Allen JW. 1980. Tissue-specific induction of sister chromatid exchanges by ethyl carbamate in mice. Environ Mol Mutagen 2(1):17–26.
[14] Zhao X, Zou H, Fu J, et al. Nitrogen regulation involved in the accumulation of urea in Saccharomyces cerevisiae.[J]. Yeast, 2013, 30(11): 437-447.
[15] Uthurry C A, Lepe J A, Lombardero J, et al. Ethyl carbamate production by selected yeasts and lactic acid bacteria in red wine[J]. Food Chemistry, 2006, 94(2): 262-270.
[16] Kobashi K, Takebe S, Sakai T, et al. Urethane-Hydrolyzing Enzyme from Citrobacter sp.[J]. Chemical & Pharmaceutical Bulletin, 1990, 38(5): 1326-1328.
[17] Tayasu I, Hirasawa R, Ogawa N O, et al. New organic reference materials for carbon- and nitrogen-stable isotope ratio measurements provided by Center for Ecological Research, Kyoto University, and Institute of Biogeosciences, Japan Agency for Marine-Earth Science and Technology[J]. Limnology, 2011, 12(3): 261-266.
[18] Wu Q, Zhao Y, Wang D, et al. Immobilized Rhodotorula mucilaginosa: A Novel Urethanase-Producing Strain for Degrading Ethyl Carbamate[J]. Applied Biochemistry and Biotechnology, 2013, 171(8): 2220-2232.
作者簡介:楊紅,女,仲愷農業工程學院在讀研究生,主要研究方向食品加工與安全。
*通訊作者:劉曉豔,女,漢族,博士,副教授
基金項目:廣式傳統發酵製品中氨基甲酸乙酯控製技術研究及其示範(B31820711)
楊紅 劉曉豔*
仲愷農業工程學院






熱點推薦
-

全十紅紅稗餅幹|中秋團圓,回家必備
-

和汪氏蜂蜜共同來普及蜂蜜結晶現象
-

拿坡海開啟西餐加盟新潮流, 大眾化家庭小西餐成未來新趨勢
-

摩購空間“霸王餐”來襲 打破傳統營銷套路帶動商家業績翻倍
-

開學季食安隱患猛增 禧雲食安開展公益行動助力意識提升
-
氨基甲酸乙酯的研究進展
-

生活飲用水微生物檢驗方法和評價標準探討
-

包裝飲用水中銅綠假單胞菌汙染情況分析
-

消毒複用餐(飲)具中大腸菌群檢測方法對比和結果分析
-

一起疑似攝入飲食感染霍亂疫情病原體的實驗室快速檢測
-
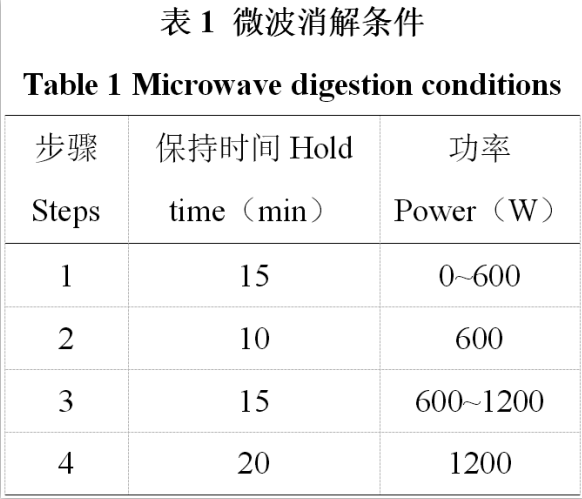
石墨爐原子吸收光譜法測定生熟茶葉中的鉛元素含量及其安全評估
-

湖北市場監管局彭明方副局長調研赤壁市磚茶質量安全監管工作
-
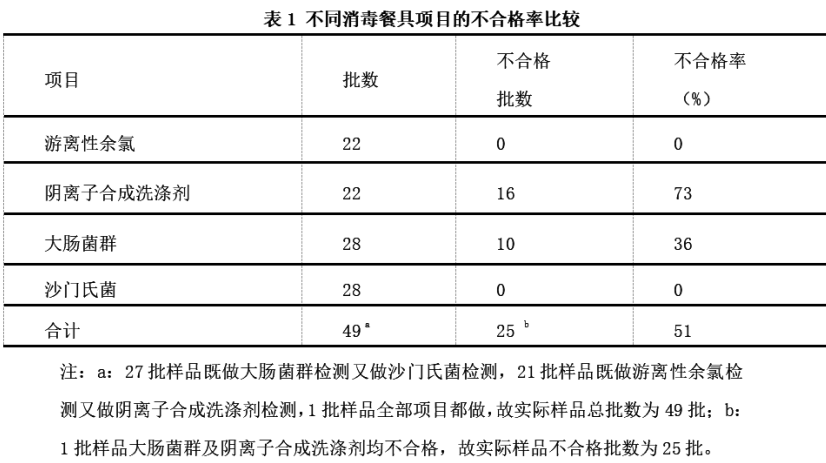
梧州市消毒餐具高風險檢出項目分析
-
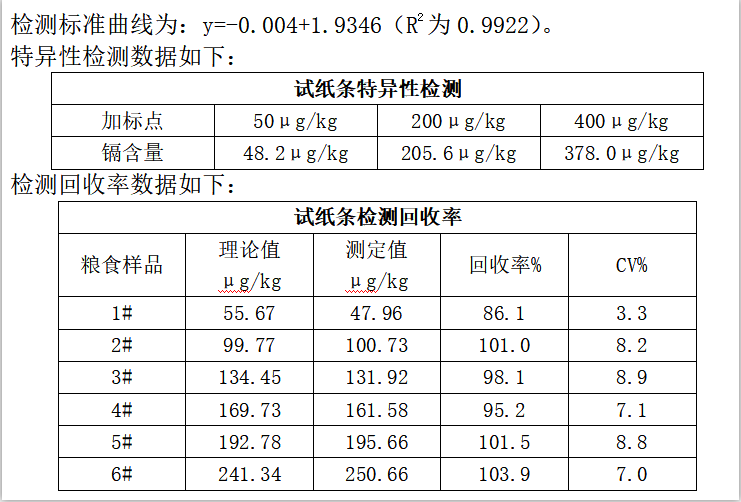
膠體金免疫層析快速定量檢測糧食中鎘殘留
-
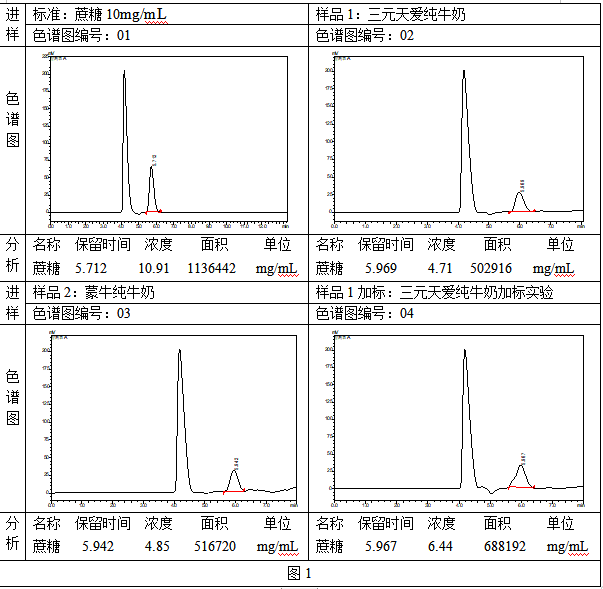
檢測乳製品中蔗糖含量的同時檢測乳糖的必要性
-
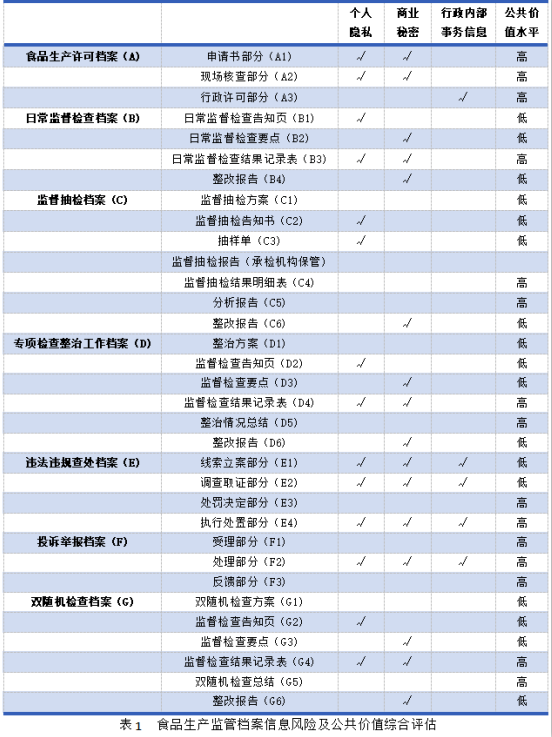
基於敏感信息保護的食品生產監管檔案開放方案研究
-

鄭州市惠濟區市場監管局召開食品生產企業世界杯賽程預測 員培訓考核會
-

餐飲具清洗消毒工藝流程對餐飲世界杯賽程預測 的重要性
-

漯河市市場監管局六舉措規範農村銷售環節世界杯賽程預測
-
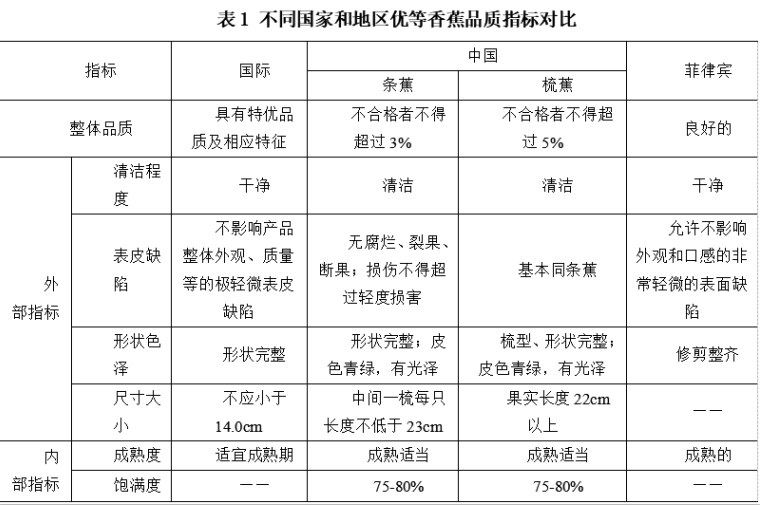
香蕉國內外評價標準對比及市場現狀分析
-
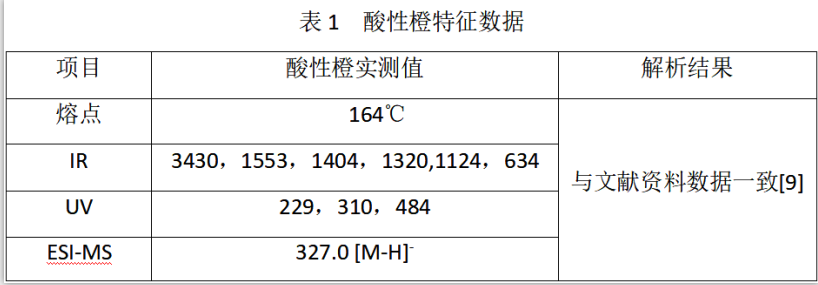
酸性橙標準品的定性分析
-

測試片方法快速檢測活性乳酸菌飲料中的乳酸菌總數
-

糧油中黃曲黴毒素檢測研究
-
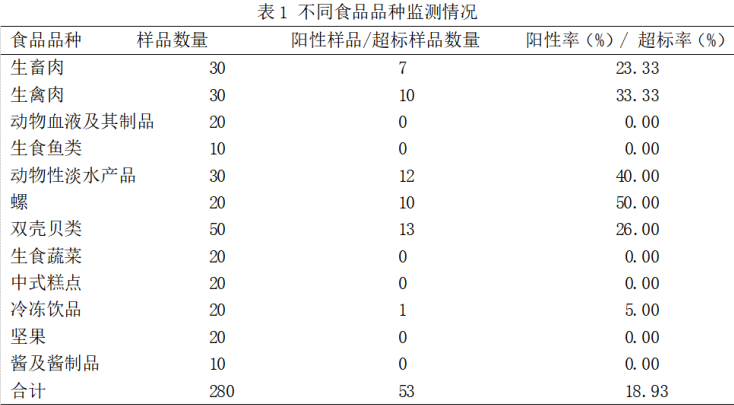
淄博市2018年食源性致病菌監測結果分析
-

重塑食物係統,根除“隱性饑餓”
-
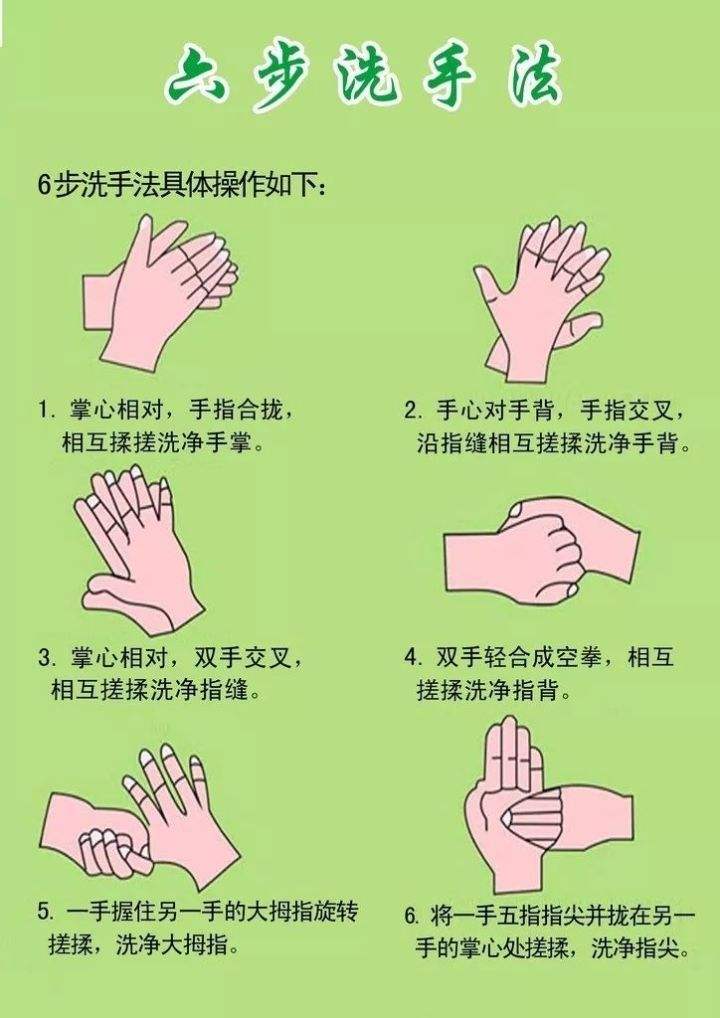
食品從業人員相關的新型冠狀病毒知識大全
-
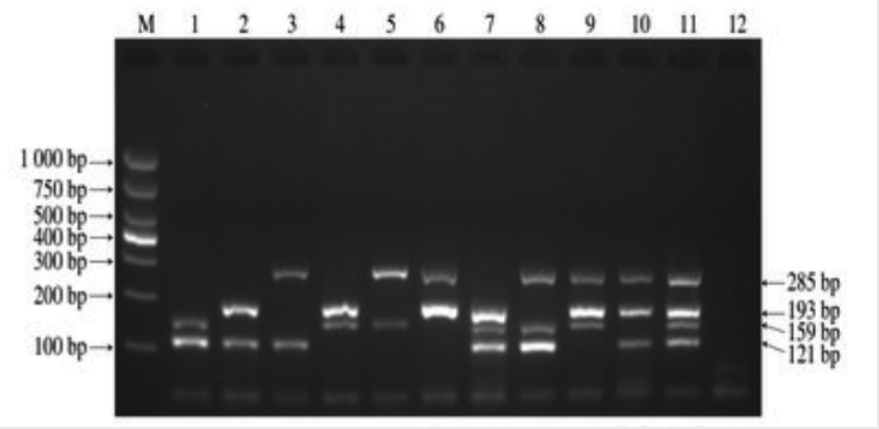
PCR技術及其在食品微生物檢測中的運用研究
-

長春市某高校教師營養知識的調查與建議
-
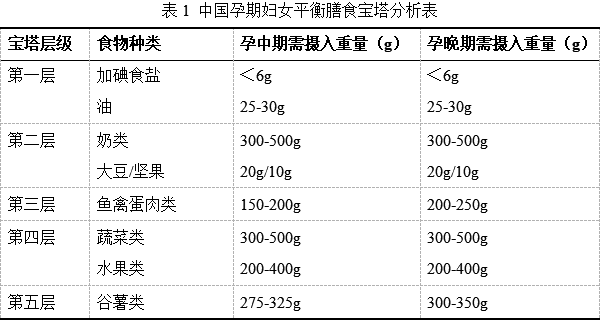
《中國孕期婦女平衡膳食寶塔》的科普解讀
-
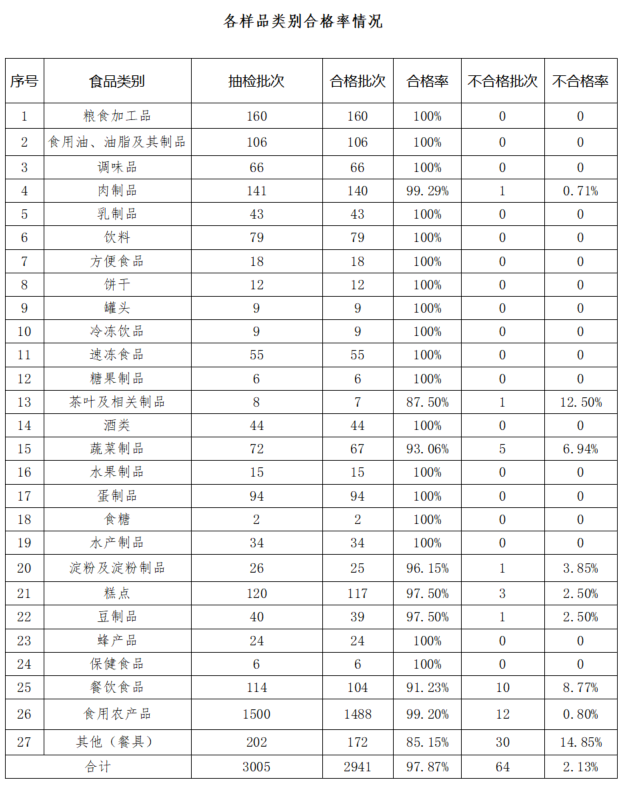
世界杯賽程預測 監督抽檢結果對日常監管工作的指導應用初探